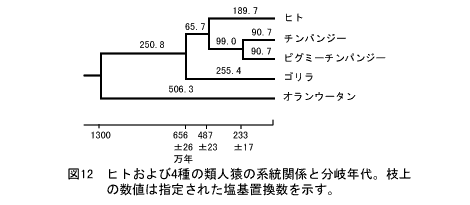
分子時計を使ったヒトとサルの分岐年代。オランウータンとの分岐が1300万年前として、他の分岐年代を求めている。
すると、ゴリラとの分岐が656万年前±26万年、チンパンジーとの分岐が467万年前±23万年前となる。
人類の起源と進化(1)も参照。国立遺伝学研究所
http://www.nig.ac.jp/museum/evolution/evolution.html
第4章 進化
生物がその形や機能を多様化させてきた過程を進化という。その結果多くの生物種が誕生し、また絶滅した。
進化という概念をはっきりとさせ、また定着化させたのはチャールズ・ダーウィン(英、1809年〜1882年)である。彼の「種の起源」(1859年)では、子は親の完全なコピーではなく、少しずつ違った形質を持つ、その中でその時代・場所の環境に適した個体はよりよく適応し、子孫を残しやすい。こうした自然選択(自然淘汰)が働くために、適応できないものはふるい落とされていく。環境が変化すれば、その環境に適応できた個体が多くいた種が残り、そうでない種は絶滅していく。こうして環境が変化すれば、自然淘汰によって適応に優れた個体群(種)が生き残っていくことになる。という考えが述べられている。
もうひとつ、あまり環境に対する適応性と関係のないような形質も、有性生殖をする動物では繁殖行動をとるときに配偶者を得られやすい(おもに雌が雄を選択)というものが発達するということもある。鳥類などで顕著であるが、雄だけが派手な飾りを持つというようなものである。ほ乳類にもそうしたものは見られる。こちらは性淘汰という。
2.木村資生の中立説
日本の木村資生(きむらもとお、1924年〜1994年)は、1968年に科学誌Natureで「中立説」を発表した。中立説とは、遺伝子の置換(分子レベルではDNAの変異=複製の際のミス)は、まわりの環境とは関係なく(その環境に有利・不利とは関係なく)、一定の割合で起こっているということである。
それまでは、ほとんどの突然変異は環境に不利であるために自然淘汰され、ごくまれに有利な突然変異が現れて進化につながるという考えであった。ところが、中立説によれば、突然変異の大部分は環境とは関係なく自然淘汰の対象にはならない、中立的なものであるということになる。
その後、木村資生の弟子である太田朋子により、変異は完全には中立ではなく、有害と中立のほぼ半分であるという「ほぼ中立説」も提唱されている。これらについては下のサイトの解説を参照。
3.分子時計
DNAなどの配列置換(変異)が、環境とは無関係にある時間で一定の割合で起こるとすれば、その置換数によって生物種の近縁関係、つまり分岐がいつころ起きたのかを推定することができる。これがDNAの分子時計である。
例えば動物のミトコンドリアDNAは1000万年で2%の置換率、またRNAの一種16SrRNAは5000万年で1%の置換率を示す。またヒトのミトコンドリアDNAの一部(Dループと呼ばれる領域の一部で500の塩基からなる部分)では、2万年で1つという置換(変異)が起こる。こうした進み方の違う分子時計をうまく組み合わせることによって、より幅広く、そして正確な時計となる。
もちろん、化石上の証拠(年代測定)を使った方法との相互比較が大切である。
分子時計を使ったヒトとサルの分岐年代。オランウータンとの分岐が1300万年前として、他の分岐年代を求めている。
すると、ゴリラとの分岐が656万年前±26万年、チンパンジーとの分岐が467万年前±23万年前となる。
人類の起源と進化(1)も参照。国立遺伝学研究所
http://www.nig.ac.jp/museum/evolution/evolution.html
 |
 |
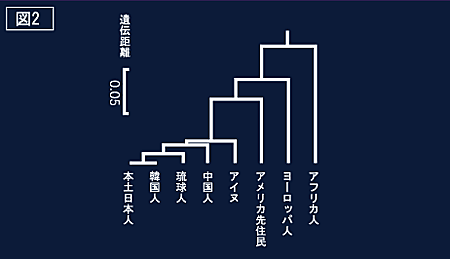
| 分子レベルで見た日本人のルーツ:国立科学博物館 http://www.kahaku.go.jp/special/past/japanese/ipix/5/5-26.html |
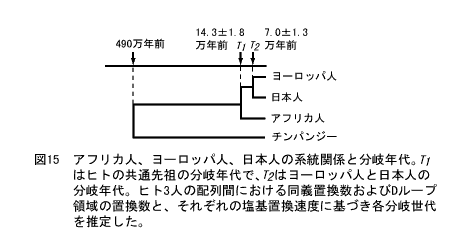
日本人のルーツ。アフリカ人との分岐が14.3万年前±1.8万年前、ヨーロッパ人との分岐が7.0万年前±1.3万年前。国立遺伝学研究所 http://www.nig.ac.jp/museum/evolution/evolution.html |
4.大絶滅
生物種の中には、子孫を残すことができずに絶滅することある。これはふつうのことである。ところが、そうした生物の絶滅が、同時にたくさんの種において起こることがある。こうした生物の大絶滅は、恐竜などが滅んだ白亜紀−古第3紀境界(K/P境界)ばかりではなく、古生代のオルドビス紀末(4億3500万年前)、デボン紀末(3億6000万年前)、二畳紀末(2億4500万年前)、中生代の三畳紀末(2億500万年前)にも起きていて、これらを5大絶滅という。
| 地質時代 | 放射年代(絶対年代) | 属レベルの絶滅率(%) | 種レベルの絶滅率(%) |
| 中生代白亜紀末 | 6500万年前 | 47 | 76±5 |
| 中生代三畳期末 | 2億500万年前 | 47 | 76±5 |
| 古生代二畳紀末 | 2億4500万年前 | 84 | 96±2 |
| 古生代デボン紀末 | 3億6000万年前 | 55 | 82±3.5 |
| 古生代オルドビス紀末 | 4億3500万年前 | 61 | 85±3 |
5大絶滅。表は「マクロ進化と全生物の系統分類」(シリーズ進化学1、岩波書店、2004年12月)のp.172より
古生代と中生代の境界の二畳紀末(P-T境界)の大絶滅がすさまじかったことがわかる。このような大絶滅の原因はよくわかっていない。地球に原因があるとする内因論としては、海水準の変動、気候変動、火山活動、あるいは巨大マントル・プルームの上昇などが考えられている。また、地球外に原因があるとする外因論としては、太陽活動の変化、われわれに近いところでの超新星爆発、小惑星(いん石)や彗星の衝突などが考えられている。中生代末の恐竜などが絶滅した事件の原因は、いん石の衝突のためであるという説が強い。
こうした大絶滅のときに運良く生き延びた生物は、それまで他の生物が占めていたい生態的な地位(ニッチェ)があいてしまうので、そこを埋めるような形で適応拡散する。こうして古生代に栄えていた両生類が大絶滅したあと、生き残った爬虫類が中生代に栄えた恐竜などの大型は忠類となって繁栄し、それらが大絶滅をしたのちほ乳類が大進化を遂げる、という図式が描ける。
こうしたことを研究したグードル(アメリカ、スティーヴン・グールド、1941年〜2002年、進化を題材とした科学解説書でも有名)は、「断続平衡説」を唱えた。生物はゆっくりゆっくり斬新的に進化するのではなく、あるときに爆発的に進化して様々な種を生み出し、その中で時代の環境に適応した種だけが生き延びて、その種はしばらく安定な時期が続くというものである。
断続平衡説に対しては批判も強い。その一つは下のサイトを参照。
進化:進化という概念そのものは、チャールズ・ダーウィン以前からもあった。例えば、チャールズ・ダーウィンの祖父エラズマス・ダーウィンもその一人である。彼は、単純な原始生物からあらゆる動物が進化してきたことを明確に述べた。ただし、自然淘汰という概念はなく、環境の変化に対する動物の反応が進化のメカニズムとした。
注目されるのはラマルク(仏、バン・バチスト・ド・ラマルク、1744年〜1829年)であろう。彼の考えは「用不用説」「獲得形質の遺伝」とまとめられる。環境の変化で必要になった器官は発達し、不用になった器官は衰えていく(用不用)。そしてその結果が子供の伝わるというものである(獲得形質の遺伝)。
ラマルクの解釈は、キリンの首が長くなったのは、高い木の葉を食べようとして一生懸命首を伸ばしたらだんだん伸びてきた、そうしたらその親から生まれた子供の首は生まれたときから長くなるいうものである。ダーウィンは、首の短いキリンは生存に不利、たまたま首が長かったキリンは生存に有利で子孫を残しやすい、そうした結果長い年月をかけてキリンは首の長い種となったという解釈となる。
当時は遺伝ということがよくわかっていなかったので、ラマルク的な考えが出るということはやむを得ないことだったと思う。しかし、20世紀に入ってからも、その亡霊が現実的な力を持つことがあった。その代表的な例がルイセンコ(旧ソ連、1898年〜1976年)一派であろう。ルイセンコは農業技師ミチューリン(旧ソ連、1855年〜1935年)の仕事(寒いロシアに適応した栽培品種をつくる研究、寒さに晒した種子は寒さに強い品種となり、それから採れる種子も寒さに強い品種になっている)を評価し、メンデルを否定した。つまり、獲得形質の遺伝を認めた。問題は、その彼が当時のソ連の最高権力者(独裁者)スターリンに取り入り、ソ連の農業・生物研究を牛耳ったことにある。つまり、ルイセンコに異議を唱えるということは、スターリンに異議を唱えるということになり、これはシベリアへの流刑、あるいは銃殺(粛清)を意味することになる。こうしてソ連の農業・生物研究は大きく遅れをとることになり、数度にわたる飢饉を招く原因にもなった。
さらにこの「ミチューリン・ルイセンコ」を信奉する人たちが、1945年〜1950年代半ばの日本にもたくさん生まれ、今から見ればお笑いの議論・論争が行われた。木村資生の師駒井卓(1886年〜1972年、京大、国立遺伝学研究所)は、反ルイセンコ派の代表的な論客である。
ルイセンコ論争が形を変えて縮小再生産されたのが、プレートテクトニクスが入ってきたころの、それに対する日本の地球科学界であっただろう。この点については、海洋出版の「月刊地球」号外No.5「地質学と地震 松田時彦教授退官記念号」(1992年4月)参照。さらに、「プレートテクトニクスの拒絶と受容」(泊次郎、東京大学出版会、2008年)も参照。
なお、進化とは生物のある種が何代にも渡って形質が変化し別の種になっていくことで、例えばポケモンのように一つの個体が変化することを指す言葉ではない(これなら変態)。ただし、宇宙の進化とか、恒星の進化、地球の進化という使い方はされている。こうした使われ方は、生物の進化学者には気になるようである。
スターリン:スターリン(1879年〜1953年)は1922年〜1953年までソ連共産党書記長(一番の権力者)、1941年〜1953年まで首相を務めた。ロシア革命(1917年2月帝政廃止、同年10月レーニンの率いるボルシェビキ(共産党)が実権を握る)が成功し、その指導者レーニン(1870年〜1924年)の死後、トロツキー(1879年〜1940年、革命ではスターリンよりも活躍したが、党内権力闘争に敗れメキシコに亡命中、スターリンが差し向けた暗殺者にピッケルを頭に打ち込まれて惨殺される)との権力闘争に勝利したスターリンは、長らく非情な独裁者として絶対的な権力をふるう。反対派(と目されたもの)は彼によって死刑(粛清)・流刑(シベリアのラーゲリ送り)に処せられた。その数は数千万人ともいわれる。
ダーウィン:チャールズ・R・ダーウィン(英、1809年〜1882年)。裕福な家庭に生まれ、大学では最初医学を学ぶが、後に牧師となる勉強をする。1831年測量船ビーグル号に博物学者として乗り組む機会を得る。これが後の彼にとって大きな影響を与えた。ビーグル号は5年をかけて世界各地を調査する。もっとも彼は船には弱く、航海中はベッドに横たわってばかりいたという。
南米やガラパゴス諸島における化石や生物の多様性を観察し、また持参したライエルの「地質学原理」も参考に、彼はそれまでのキリスト教の天変地異説に疑問を持つようになり、進化という概念を持つに至る。また、マルサスの「人口論」や、ハトの人為的な品種改良もヒントになったという。
航海から帰還後、働く必要がない(ほど裕福だった)ダーウィンは、思索の生活に入る。そして1858年にウォーレスとの共同論文の形で考えを発表し、翌1859年に「種の起源」を出版する。本は即日完売だったという。
当然反発も強く、とくにキリスト教にとっては、進化論は神の創造を否定するものとして激しい批判を行った。現在になっても、いわゆるキリスト教原理主義(聖書に書いてあることはすべて正しいという宗派)の強いアメリカのいくつかの州では、たびたび学校では進化論を教えてはいけないという運動が起こる。
「種の起源」が出版された翌年(1860年)のオクスフォードにおける公開討論会でのキリスト教の指導者ウィルバーフォース主教の質問、「…貴方の祖先がサルというなら、それは母方ですか、父方ですか。」に対し、ダーウィン派のハクスリー(自らダーウィンの番犬と自称していた)は、「私が卑しいサルを祖父にもちたいのか、それとも、すぐれた才能と大きな影響力を科学的議論をからかうためにだけに用いる人間を祖父にもちたいのかという質問なら、私はためらうことなくサルを選びます。」と答えたという。
また、当時はまだ遺伝ということがよくわかっていない時代だったので、なぜ変異が次の世代に伝えられるのかという批判も受けた。これが解決するのは、メンデルによる遺伝の法則の発見以降ということになる。
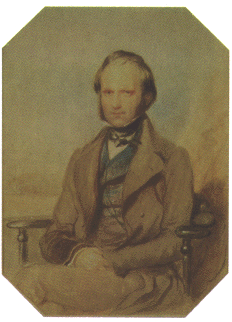 |
 |
 |
| 若いころのダーウィン | 中年のダーウィン(1859年) | 晩年のダーウィン |
| 写真はいずれもhttp://www.mun.ca/biology/scarr/Darwin.htm | ||
 |
 |
| ビーグル号 http://www.victory-cruises.com/beagle_wulaia.html |
ビーグル号の航海 http://www.portalciencia.net/geniosdarwin.html |
ウォーレス:アルフレッド・R・ウォーレス(英、1823年〜1913年、博物学者)。アマゾンや東南アジアの動物相を研究し、とくにカリマンタン(ボルネオ)とスラウェシ(セレベス)島を境界として、動物相に根本的な相違があることを認めた(ウォーレス線の提唱)。こうした経験から、ウォーレスもダーウィン同様の自然淘汰による進化という概念を持つに至った。彼の手紙を受け取ったダーウィンは、その考え方のあまりの一致に驚いたという。
ダーウィンは自分より若いウォーレスを気遣い、またウォーレスもダーウィンを尊敬していたようである。ただし、ヒト(人類)も動物の一員と見るダーウィンと、ヒト(人類)は別格と見るウォーレスは、この点で大きく違っていた。じっさい、ウォーレスは晩年は心霊現象の研究に没頭してしまう。
マルサス:トーマス・R・マルサス(英、1766年〜1834年、経済学者)。「人口論」は1798年。その中で、人口の増加は、つねに食糧の増産を上回る傾向にあることを述べた。食糧が増産傾向にあれば、人口はそれを刺激としてさらに増加し、逆に食糧の増産が人口の増加に見合わないと、飢饉、疫病、さらには戦争によって人口の増加が抑制されるとした。
ハクスリー:トーマス・H・ハクスリー(英、1825年〜1895年、生物学者)。自らダーウィンの番犬(ブルドッグ)と称した。難しい進化という概念を、一般の人にもわかりやすい形にかみ砕いた講演を行い、進化論を広めることに大きな役割を果たした。現在必要とされる、科学者と一般の人との間を取り持つインタープリーター(翻訳者)の先駆けともいえる。孫のジュリアン・S・ハクスリー(英、1887年〜1975年)は祖父と同じく有名な生物学者であり、これまた祖父と同じ優れた科学解説家でもある。その弟のアルダス・L・ハクスリー(英、1894年〜1963年)は、小説家・文明批評家。著書に(逆)ユートピア世界を描いた「すばらしい新世界」がある。
メンデル:G・J・メンデル(オーストリア、1822年〜1884年)は農家に生まれ、修道士となった。土地の名産ワイン用のブドウを改良しようとして、まずエンドウマメを用いて修道院の庭で遺伝の研究を行った。そして、対立する形質、例えば豆が黄色と緑色、形が丸いとしわが寄っているという形質が、どのように子孫に伝わるかを調べた。その結果、そうした形質には子孫に現れやすい優性と現れにくい劣性があり、黄色が優性、緑色が劣性、丸い形が優性、しわが寄るものが劣性ということがわかった。そして、その現れ方は優性:劣性が3:1という割合になること、またそれぞれの形質は違った形質とは独立であることも明らかにした。
例えば、黄色で丸い豆を付けるエンドウと、緑色でしわのある豆を付けるエンドウを掛け合わせると、できる豆は黄色で丸い:黄色でしわ:緑で丸い:緑でしわが9:3:3:1になることを明らかにした。
メンデルが明らかにした遺伝に関する法則をメンデルの法則(優劣の法則、分離の法則、独立の法則の3つ、詳しくは下のサイトの解説を参照)という。
彼はこうした研究を1865年に発表(刊行は1866年)したが、その後34年間無視されてしまった。彼の仕事が再評価されたのは、1900年に、ド・フリース(オランダ、1848年〜1935年)たち3人が独立にメンデルの法則を再発見したことによって、ようやく34年前のメンデルの論文が明らかになり、メンデルが評価されるようになった。
遺伝をつかさどる物質が染色体に含まれる遺伝子であり、その実体はDNAである。真核生物においては、遺伝子は細胞の核の中に入っている。
なお、優性・劣性とは、ある形質があくまでも子孫に現れやすいか現れにくいかということであり、生物学的・社会学的に優れた性質・劣った性質ということではない。同じ発音なので、よく混同されるのが優生、あるいは優生学である。優生とは優ている(と誰かが恣意的に判断した)性質のことである。そうした親から生まれる子供も優れている、そうでない親から生まれた子供は劣っている、だから優れた親はたくさん子孫を残すべき、逆に極端には劣った親は子供を残してはいけないとするもの(運動)である。ヒトラー(ドイツ、1889年〜1945年)が率いるナチス(ナチ党)が行ったユダヤ人に対するホロコースト(大量虐殺)の根拠にもなった。すなわち、優れたアーリア人(ドイツ人)だけが残るべきで、劣ったユダヤ人は殺して根絶やしにすべきだという考えであった。今日でも、こうした優生学的な考えを持っている「知識人」がいるので注意しなくてはならない。あの木村資生氏もそういう傾向がある。
 |
 |
| メンデル http://www.library.villanova.edu/ services/exhibits/gregor_johann_mendel.htm |
メンデルが研究に使ったブドウ。東京の小石川植物園(東京大学理学系研究科附属植物園)に植えられている。説明版には「メンデル葡萄:このブドウは遺伝学の祖ヨハン・グレゴール・メンデルが実験に供したものの分株である。大正2年(1913)に三好学教授がチェコスロバキアのブリュン(現在のブルノー)にメンデルの遺跡を訪ねた時、旧農園にあったブドウの分譲を依頼し、その翌年に送り届けられたものである。」とある。2001年10月撮影。 |
木村資生(きむらもとお):木村資生(1924年〜1994年)は愛知県に生まれ、京都大学で植物学を修める。長らく国立遺伝学研究所に勤める(一時アメリカで研究生活を送る)。1968年に発表した中立説で世界に衝撃を与える。1976年に文化勲章を受ける。

木村資生と彼の論文が載ったNature:国立遺伝学研究所
http://www.nig.ac.jp/museum/evolution/evolution.html
はじめての進化論(河田雅圭):http://meme.biology.tohoku.ac.jp/INTROEVOL/index.html
国立遺伝学研究所遺伝学電子博物館:http://www.nig.ac.jp/museum/msg.html